
NATURAL REGENERATION CAPACITY OF VITELLARIA PARADOXA (SHEA TREE) ON THE HILLS OF BASSAR (NORTH-TOGO)
Dermane Wagbe
Tchapo 1 ![]() , Djangbedja Minkilabe 1
, Djangbedja Minkilabe 1![]() , Kamou
Hodabalo 2
, Kamou
Hodabalo 2![]()
1 Laboratory
of Biogeographic Research and Environmental Studies (LaRBE), University of Lome,
Togo
2 Laboratory of Botany and Ecology
Vegetal (LBEV), University of Lome, Togo
|
|
|
ABSTRACT |
|
|
As the
consequence of anthropic pressure along with climatic factors, the vegetation
of the hills of Bassar is degrading day after day and a regression of many
woody species is observed. Vitellaria paradoxa is a highly valued
multi-use species, but it is threatened with extinction in the area. The
"bell" and "L" shapes of the diameter class distribution
histograms reflect aging and/or disturbed populations. Stand disturbance is
illustrated by the low proportion of large diameter class individuals. The
"L" shape reveals an advanced state of degradation characterized by
a near absence of large diameter trees. The study of the natural regeneration
of Vitellaria
paradoxa on the hills of Bassar shows that the
density of regeneration by seedlings, shoots and suckers depend on the
diameter of the individual stem. The larger the diameter increases, the
higher the density of seedlings becomes. However, plantlets stemming from
seedlings are higher, and weaker from suckers and shoots. The density of Vitellaria paradoxa seedlings is lower in shrub savannahs than in tree savannahs. The
analysis of variance showed that whether under the crown or outside the
crown, there was a significant difference between individuals from regeneration
in all the formations studied. In the inventory plots, the densities of young
trees are equally variable under the crown and outside the crown as those of
large trees. |
|||
|
Received 11 August 2022 Accepted 10 September 2022 Published 26 September 2022 Corresponding Author Dermane
Wagbe Tchapo, dermanetchapo@gmail.com
DOI10.29121/granthaalayah.v10.i9.2022.4764 Funding: This research
received no specific grant from any funding agency in the public, commercial,
or not-for-profit sectors. Copyright: © 2022 The
Author(s). This work is licensed under a Creative Commons
Attribution 4.0 International License. With the
license CC-BY, authors retain the copyright, allowing anyone to download,
reuse, re-print, modify, distribute, and/or copy their contribution. The work
must be properly attributed to its author.
|
|||
|
Keywords: Hills of Bassar, Natural Regeneration,
Capacity |
|||
1. INTRODUCTION
The natural regeneration of forest resources is a complex process that is controlled by several biological, ecological, demographic, and historical factors Bucci and Borghetti (1997). In the same line, ecologists are interested in the regeneration of woody species in the process of ecosystem renewal. In the plant succession dynamics, the regrowth of plants contributes to the maintenance of the specific diversity of woody species and the structure of future vegetation is drawn from the existing young tree populations Maingi and Marsh (2006).
Studies on regeneration in the context of tree population dynamics in West Africa have grown over the last decade Sinsin et al. (2004), Gaoué and Ticktin (2007), Gaoué and Ticktin (2009), Koulibaly et al. (2010). According to these studies, the tree distribution pattern and their regeneration is affected by many biotic and abiotic factors and their interactions. Not only is regeneration capacity favored by an optimal density of seed-bearing individuals but also by facilitating relationships Sinsin et al. (2004), Biaou et al. (2011). Studies carried out on the demography of the seedlings Ouédraogo et al. (2006), Ouédraogo et al. (2009), Avocèvou-Ayisso et al. (2009), regeneration mechanisms Ky-Dembele et al. (2007) and the impact of influence on the structure of the vegetation Bondé et al. (2013), Avocèvou-Ayisso (2009), Schumann et al. (2011), Nacoulma e al. (2011) have also highlighted the determining role of the soil in tree establishment and regeneration.
Studies dealing with the effect of topography on regeneration mainly concern temperate mountain forests Coop et al. (2010), Robert (2003). They show the negative effect of elevation and slope on forest regeneration, and the positive effect of gaps on the diameter and height structure of populations Coop et al. (2010), Robert (2003). But in the context of monitoring vegetation in the Sudanian area, which is particularly exposed to deforestation Mama et al. (2014), the issue of the relationship between the relief and the regeneration capacity (the number of regenerating species and the density of regeneration) has not yet been dealt with.
Regeneration capacity is necessary for an ecosystem to have good plant dynamics. Even if the stock of seeds, the germination capacity of each seed and the modes of dispersion are determinant factors of this capacity, the edaphic conditions are nonetheless essential for regeneration Ouédraogo et al. (2009). Human activities have been observed on the hills of Bassar and its surroundings. For local populations, "shea trees (Vitellaria paradoxa) are more used for charcoal." Moreover, the plant species of the study environment and based on the species listed on the red list of the IUCN in Togo, the objective of this research is to assess the natural regeneration capacity of Vitellaria paradoxa on the hills of Bassar and its surroundings.
2. Study area
The study was conducted on the hills of Bassar and its surroundings in the prefecture of Bassar. Bassar is located in the northern part of Togo in the southwest of the Kara region. It is on Longitude 0°15’ and 1°30’ E and Latitude 9°25’ and 10°10’ N. The prefecture of Bassar is limited to the North and North-West by the prefecture of Dankpen, to the North-East by the Prefectures of Kozah and Doufelgou, to the East by the Prefecture of Assoli; to the South-East by the Prefecture of Tchaoudjo, to the South by the Prefecture of Sotouboua and to the West by the Republic of Ghana. As presented, the study site is located in a hilly environment. Figure 1
Figure 1
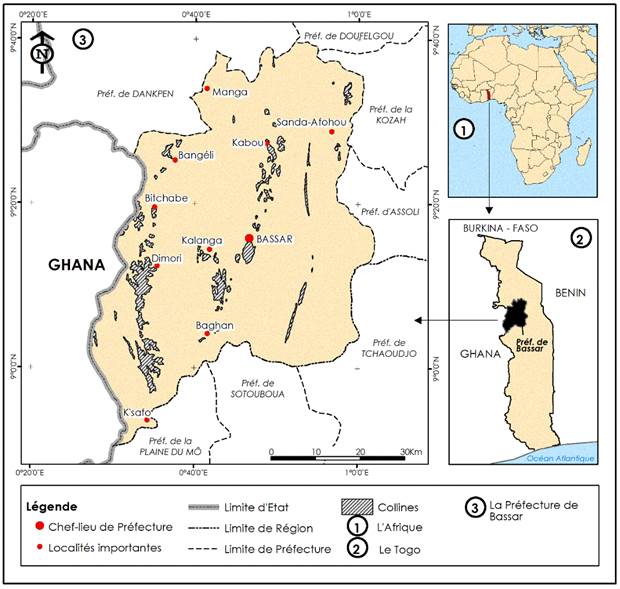
|
Figure 1 Map of the Study Area Source ADDRA C. T.,
Togo Atlas Map (1984), Updated Map (2022) |
The landscape of the prefecture of Bassar is made up of two large units which are divided between the north and the south. It is dominated by hills and plateaus oriented North-South and characterized by a succession of anticlines and synclines with breaks and overlaps related to the brittle nature of the geological formations Gnon (1967), Potapou (1997). The north is the domain of a plain landscape which is the extension of the Oti plain. It is a monotonous plain. The altitudes vary between 100 and 200 m and drop slightly towards the Oti River. In opposition to the large plain of the north is a mountainous area in the south where Bassar is located. This hilly area has north-south two subset orientation. They are separated by a central depression at an altitude of between 200 and 300 meters and about 25 to 30 kilometers wide. The site is drained by the Katcha River Gnon (1967), Potapou (1997).
According to the ecological subdivisions of Togo Ern (1979), the prefecture of Bassar is located between ecological zones I and II. The plant formations are varied according to the topographic forms. Indeed, the slopes of the mountain are covered with tropical sparse forests while the plains areas are the domain of wooded savannah. The main plant species are: néré (Parkia biglobosa), shea (Vitellaria paradoxa), baobab (Adansonia digitata), kapok tree (Bombax costatum), cheese tree (Ceiba pentandra), teak (Tectona grandis), palm oil. (Elaeis guineensis), cailcedrat (Khaya senegalensis), iroko (Milicia excelsa), false iroko (Antiaris africana), lingue (Afzelia africana), Isobernia doka and ebony (Pterocarpus erinaceus). These plant formations develop on a diversity of soils, the most represented of which are leached tropical ferruginous soils, poorly evolved soils from erosion, ferralitic soils and vertisols.
The prefecture of Bassar has an area of 3620 km², approximately 6.4% of the total area of Togo and 31.12% of the Kara Region. It is populated by19,717 inhabitants in 2010 RGPH4, (2010), for a density of 33 inhabitants per km². Located about 400 kilometers from the sea, the prefecture of Bassar is made up of a mosaic of ethnic groups such as Ncam speakers, Fulfilde speakers, Konkomba speakers, Kabyè speakers, Nawdm speakers, Tem speakers, Lamba speakers, Yorouba speakers and Hausa speakers. Ncam is the original and most widely spoken language in the area.
The main economic activity is agriculture. It employs 90% of the active population. Another activity is the extensive breeding of cattle and goats. The native Ncam ethnic group of the region have known iron metallurgy since the 19th century. About 700 blast furnaces can be numbered on the various extraction sites of Bandjeli and Nangbani. The development of this metallurgy had harmful consequences on the vegetation. Indeed, it uses a considerable amount of charcoal to melt the iron.
3. MATERIALS AND METHODS
3.1. MATERIALS
This research required the use of the following equipment and instruments:
· a GPS to take the geographical coordinates of plots.
· a compass for orientation.
· a pentadecameter for sizing plots.
· a cutter for installing plot corner stakes.
· a digital camera for taking pictures.
3.2. METHODOLOGY
3.2.1. Data Collection
In 50 m x 50 m plots, every Vitellaria paradoxa individual whose diameter is less than or equal to 10 cm were taken into account in the study of regeneration. Diameters greater than 10 cm were measured at 1.30 m from the ground using a tape measure. The regeneration study was carried out in the 16 m² (4 m x 4 m) sub-plots in the wooded savannahs, shrub savannahs and open forests. In the study, every young plant whose diameter is less than or equal to 10 cm are considered as young shoots. These young shoots were identified taking into account their abundance and their origin (seedlings, shoots, and suckers). Thus, under the foot of an individual, small plots of 4 m x 4 m were made, and all these young shoots were systematically counted.
A total of 90 plants were taken into account in this study adopted the Ræbild et al. (2012) method. This method was then modified according to the ecological and topographical conditions of the study area. Under the crown and outside the crown cover of each individual of the species, a square plot of 16 m² (4 m x 4 m) was made. First subplots were placed under and at the edge of the crown while second subplots were placed completely outside the crown cover Figure 2. Selected trees were painted, and plot boundaries were semi-permanently marked with red/white ribbons in each corner. In case the red/white ribbons were removed in the second season, the plot could be established since the painting on the tree indicated the direction of the plot.
The plots were assessed monthly from August 10, 2020, to October 31, 2021. Throughout the fieldwork, informal conversations with local people were carried out in order to obtain changes in population density at the level of the species over time.
Figure 2
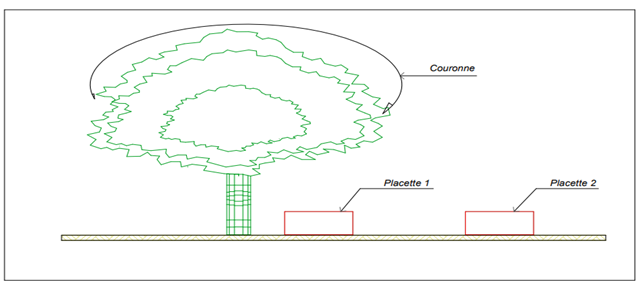
|
Figure 2 Positioning of the Plots in Relation to the Tree Source Fieldwork,
2021 |
3.2.2. Statistical analyses
Densities were analysed by Analysis of Variance and using a model including the effects of the area and the environment. The model including the effects of area is land use and the interaction between area and land use. It was necessary to use a logarithmic transformation to obtain residuals according to a normal distribution.
Trees were subdivided into 10 cm diameter classes, and the size class distributions are presented graphically. The number of seedlings, shoots and suckers in the regeneration study was analysed for each type of seedlings, shoots, and suckers. However, the numbers of seedlings under and outside the crowns seemed to be done according to different distributions.
Due to the fact that some seedlings have been grazed, burned, or cut were recorded the analysis of these parameters were mentioned. This problem could not be solved by transformations and made it impossible to perform Analysis of Variance with the effects of the main factors (area, land use, species, under crown, out of crown) without violating the assumptions of homogeneity of the variance and of the normal distribution of the deposits. Instead, the analyses were performed in two stages. First, the differences between the number of seedlings under the crown and outside the crown were analysed using the Pearson test, this test is valid even in cases where the data does not follow a normal distribution. These analyses were carried out for each combination. The second step was to analyse the number of seedlings by general linear models in order to analyse the differences between areas, land use and number of seedlings.
4. RESULTS
4.1. DENSITY AND DIAMETER CLASS DISTRIBUTIONS ACCORDING TO THE REGENERATION
The average diameter of Vitellaria paradoxa is 23 cm ± 2.75 in the wooded savannahs with many individuals from 10 to 40 cm Figure 3.
Figure 3
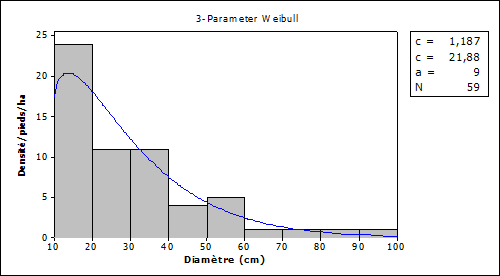
|
Figure 3 Density Distribution and Diameter Classes According to the Regeneration in Wooded Savannah Areas Source Fieldwork,
2021 |
In shrubby savannahs, the average diameter of Vitellaria paradoxa individuals is 21 cm ± 1.4 with many having a diameter of between 10 and 20 cm Figure 4.
Figure 4
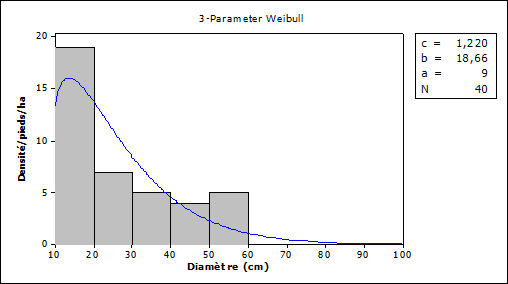
|
Figure 4 Density Distribution
and Diameter Classes According to Regeneration in Shrubby Savannah Areas Source Fieldwork,
2021 |
On the other hand, in open forests, the average diameter of individuals of Vitellaria paradoxa is 30 cm ± 2.1. Diameter Individuals between 20 and 40 cm are more numerous Figure 5.
Figure 5
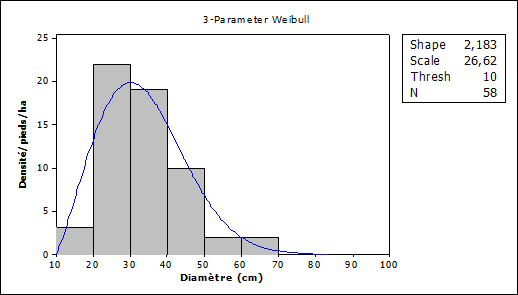
|
Figure 5 Density Distribution and Diameter Classes Following Regeneration in Open Source Fieldwork,
2021 |
4.2. NATURAL REGENERATION DYNAMICS IN DIFFERENT PLANT FORMATIONS
4.2.1. Natural regeneration Dynamics in woody savannahs
Vitellaria paradoxa young plants are constantly present. The Analysis of Variance showed that both under the crown and outside the crown, there is a significant difference between the individuals resulting from the regeneration in all the formations that have been investigated. In the inventory plots, the densities of young plants are just as variable both under the crown and outside the crown as those of large trees Table 1. The regeneration of Vitellaria paradoxa happened more exclusively by sowing (10.88 ± 1.00) followed by regeneration by suckers (5.19 ± 0.44). On the other hand, regeneration is weak by suckering Table 1. Underground woody tubers are often several years old, but young plants from sprouts are mostly seasonal.
Table 1
|
Table 1 Seedling Average Density and Regeneration Origin in Wooded Savannahs |
||
|
Under the Crown |
Outside the Crown |
|
|
Density (number of
seedlings per plot of 4 m x 4 m) |
6.23 ± 2.02 |
3.09 ± 0.001 |
|
Seedlings grown from new
seed germinations (%) |
10.88 ± 1.00 |
5.78 ± 0.09 |
|
Shoots (%) |
5.19 ± 0.44 |
3.24 ± 0.07 |
|
Suckers (%) |
3.14 ± 0.15 |
1.2 ± 0.012 |
|
Source Fieldwork, 2021 |
||
4.2.2. Dynamics of natural regeneration in shrubby savannahs
The density of Vitellaria paradoxa seedlings is lower in shrubby savannahs than in woody savannahs. The Analysis of Variance showed that both under the crown or outside the crown, there is a significant difference between the individuals resulting from the regeneration in all the formations that have been investigated. In the inventory plots, the densities of young plants are just as variable both under the crown and outside the crown as those of large trees Table 2. The regeneration of Vitellaria paradoxa took place more exclusively by sowing (8.05 ± 1.20) followed by regeneration by suckers (3.17 ± 0.24). On the other hand, regeneration by suckering is weak Table 2. Underground woody tubers are often several years old, but young plants from shoots are mostly seasonal.
Table 2
|
Table 2 Average Density of Seedlings and Origin of Regeneration in Shrubby Savannahs |
||
|
Under the Crown |
Outside the Crown |
|
|
Density (number of
seedlings per plot of 4 m x 4 m) |
5.12 ± 2.00 |
2.2 ± 0.001 |
|
Seedlings grown from new
seed germinations (%) |
8.05 ± 1.20 |
4.45 ± 1.00 |
|
Shoots (%) |
3.17 ± 0.24 |
1.54± 0.17 |
|
Suckers (%) |
1.55 ± 0.05 |
1.03 ± 0.04 |
|
Source Fieldwork, 2021 |
||
4.2.3. Dynamics of natural regeneration in open forests
Young plants are constantly present in Vitellaria paradoxa population. The Analysis of Variance showed both under the crown and outside the crown, there is a significant difference between the individuals resulting from the regeneration in all the formations that have been investigated. In the inventory plots, the densities of young plants are just as variable both under the crown and outside the crown as those of large trees Table 3. The regeneration of Vitellaria paradoxa takes place more exclusively by sowing followed by suckering regeneration. On the other hand, regeneration is weak by suckering Table 3.
Table 3
|
Table 3 Seedling Average Density and Origin of Regeneration in Open Forests |
||
|
|
Under the Crown |
Outside the Crown |
|
Density (number of seedlings
per plot of 4 m x 4 m) |
12.43 ± 3.01 |
7.28 ± 1.82 |
|
Seedlings grown from new
seed germinations (%) |
20.00 ± 3.00 |
9.17 ± 2.30 |
|
Shoots (%) |
11.80 ± 2.70 |
5.87 ± 1.96 |
|
Suckers (%) |
5.74 ± 1.28 |
2.22 ± 0.37 |
|
Source Fieldwork, 2021 |
||
4.3. DIAMETER CLASS REGENERATION IN VITELLARIA PARADOXA
The demographic distribution in diameter classes in Vitellaria paradoxa is illustrated by histograms Figure 6. The density of regeneration by seedlings, shoots and suckers depends on the diameter of the foot of the original individual. The larger the diameter increases, the higher the density of seedlings becomes. However, seedlings from sowings are higher, and weaker by shoots and suckers.
Figure 6
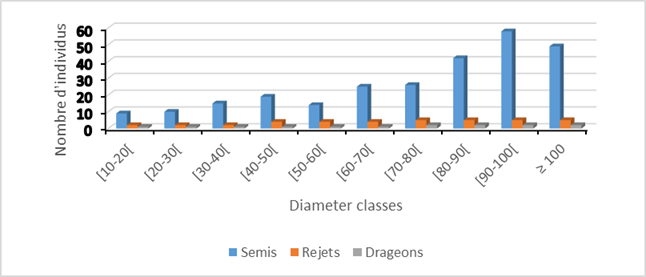
|
Figure 6 Diameter Class Regeneration Source Fieldwork,
2021 |
The following plates show the different types of natural regeneration mode encountered in the field in Vitellaria paradoxa.
Figure 7

|
Figure 7 Shoot Regeneration Source Authors,
2021 |
Figure 8

|
Figure 8 Sowing Regeneration Source Authors,
2021 |
Figure 9

|
Figure 9 Seeds of Vitellaria Paradoxa on the Research Area Source Authors,
2021 |
5. DISCUSSION
The "bell" and "L" shapes of the distribution histograms in diameter classes reflect aging and/or disturbed populations. Tree population disturbance is illustrated by the low proportion of individuals in large diameter classes. The “L” shape reveals an advanced state of degradation that is characterized by a virtual absence of large diameter trees. The natural regression factors are mainly endogenous and climatic, but these species are also subject to strong anthropogenic pressure which may be reflected in the heterogeneity of diameters among populations. The seasonal renewal of stems by shoots of underground woody tubers of Vitellaria paradoxa is similar to vegetative reproduction. The dominance of the survival dynamics or a state of overcoming crossbreed reproduction capacities of seed trees. According to Bellefontaine et al. (2000), vegetative regeneration often appears as an adaptation strategy to disturbances and climatic hazards in crossbreed species. It is not favourable to the conservation and maintenance of plant populations because of genetic erosion. Moreover, natural dissemination by vegetative means does not ensure spatial propagation of plants on a scale as large as seeds. A natural dynamic in favor of the vegetative mode of reproduction can reflect increased environmental pressure on woody plants. In the natural environment, seedlings have to face drought, bush fires and predators. Very few of them survive the combination of these three factors. The survival of some seedlings is possible thanks to suckers or their own ability to develop a woody underground tuber. The loss of the above-ground parts, often by drying out in the dry season or grazed in the wet season, means that the seedlings are in a perpetual dynamic of "restarting growth". In the current context, sexual reproduction seems to be a combined action of drought, fires, and predators (cattle, insects, rodents, fungi, etc.). Vitellaria paradoxa has a good capacity of vegetative regeneration by seed in unsuitable conditions, especially regarding to drier climatic conditions and increasingly accentuated anthropogenic pressure.
Vitellaria paradoxa trees are present everywhere in the ecosystems that have been visited. For a layman, (including ourselves), the tree population seems to be of old age because there is an absence of small trees. However, the current data show an abundant regeneration especially in fallows where there exists many trees of small diameter classes. Seedlings of Vitellaria paradoxa are relatively common away from trees and the number of seedlings per hectare is estimated to an average field of at least 15-25 in all ecosystems. However, as shown by the diameter distribution, seedlings do not grow into trees, most likely because they are removed when farmers go in search of wood for energy purposes. This phenomenon is confirmed by the sudden decrease in the number of seedlings in shrub and tree savannahs. In open forests, on the contrary, seedlings can develop into trees without restrictions. This is related to the topographic situation of these forest ecosystems. These results are contrary to those obtained by Kelly et al. (2004) who found that Vitallaria paradoxa regenerated mainly in fallows. Comparable levels were found in shrubby savannahs and in tree savannahs. The results of Pare et al. (2009) in Burkina Faso showed a high proportion of seedlings of Vitallaria paradoxa in the forests of the Sudanian zone of Burkina Faso. The number of seedlings of Vitellaria paradoxa was almost significantly higher under the crowns than outside. It would be natural to assume that seedlings arise from seeds either falling from the parent tree above, or from seeds brought to the tree by a vector - birds or bats that sit on the tree and eat the fruit of Vitallaria paradoxa or by human beings who take their break under the tree and also eat fruit. Indeed, farmers or hunters often eat the fruits of Vitallaria paradoxa during the ''hunger period'' at the beginning of the rainy season, especially during farm activities. The fact that Vitellaria paradoxa nuts are also common under other trees shows that the seeds are indeed dispersed. Kelly et al. (2004) found that Vitellaria paradoxa trees tended to cluster with increasing aggregation from fields to fallows and forests. Under a Vitallaria paradoxa, the number of seedlings shows a clear variation with the seasons. It sometimex reach a peak during the rainy season. In Uganda, Okullo et al. (2004)observed similar results in most fruit trees. However, this phenomenon is not clear, but could be due to the emergence of root suckers from destroyed seedlings Ky-Dembele et al. (2007). According to Hall et al. (1996) and Bellefontaine (2005), Vitellaria paradoxa can survive repeatedly after the removal of above-ground parts by fire. As a consequence, it can germinate again the following rainy season. The decline in the number of seedlings can be explained by drought, but grazing by animals and cutting by humans may be some other reasons
6. CONCLUSION
The natural regeneration of tree population is a complex process that is controlled by several factors. From this study, Vitelaria paradoxa seedlings are constantly present. The density of regeneration by seedlings, shoots and suckers depends on the stem diameter of the original individual. The larger the diameter increases, the higher the density of seedlings becomes. However, seedlings from shoots are higher, and weaker by shoots and suckers. Domesticating the plant is very favorable. For this purpose, the knowledge of the usefulness of Vitellaria paradoxa (shea) would be important to develop projects for planting the species on the hillsides of Bassar.
CONFLICT OF INTERESTS
None.
ACKNOWLEDGMENTS
None.
REFERENCES
Avocèvou-Ayisso, C., Sinsin, B., Adégbidi, A., Dossou, G., and Van Damme, P., (2009). Sustainable Use of Non-Timber Forest Products : Impact of Fruit Harvesting on Pentadesma Butyracea Regeneration and Financial Analysis of its Products Trade in Benin, Forest Ecology and Management, 257(8), 1930-1938. https://doi.org/10.1016/j.foreco.2009.01.043
Bellefontaine, R., (2005). Pour de nombreux ligneux la reproduction sexuée n'est pas la seule voie : analyse de 875 cas. Sécheresse, 16, 317-328.
Bellefontaine, R., Edelin, C., Ichaou, A., Du Laurens, D., Monsarrat, A., Loquai, C., (2000). Le drageonnage, alternative aux semis et aux plantations des ligneux dans les zones semi-arides : protocole de recherches. Sécheresse, 4(11), 221-226.
Biaou, S.S.H, Holmgren, M, Sterck, F.J., and Mohren G.M.J., (2011). Stress-Driven Changes in the Strength of Facilitation on Tree Seedling Establishment in West African Woodlands. Biotropica 43, 23-30. https://doi.org/10.1111/j.1744-7429.2010.00642.x
Bondé, L., Ouédraogo, O., Kagambega, F., and Boussin, I. J., (2013). Impact des gradients topographique et anthropique sur la diversité des formations ligneuses soudaniennes. Ciorad, 318(4), 15-26. https://doi.org/10.19182/bft2013.318.a20514
Bucci, G., Borghetti M., (1997). Understory Vegetation as a Useful Predictor of Natural Regeneration and Canopy Dynamics in Pinus Sylvestris Forests in Italy. 18(4), 485-501. https://doi.org/10.1016/S1146-609X(97)80036-7
Coop, J. D., Massatti, R. T., and Schoettle, A. W., (2010). Subalpine Vegetation Pattern Three Decades After Stand-Replacing Fire: Effects of Landscape Context and Topography on Plant Com- Munity Composition, Tree Regeneration, and Diversity. Journal of Vegetation Science, 21 (3), 472-487. https://doi.org/10.1111/j.1654-1103.2009.01154.x
Ern, H., (1979). Die Vegetation Togo, Gliederrung, Gfährdung, Erhaltung, Willdenowia, 9, 295-312.
Gaoué, O. G., Ticktin T., (2007). Patterns of Harvesting Foliage and Bark from the Multipurpose Tree Khaya Senegalensis in Benin : Variation Across Ecological Regions and its Impacts One Population Structure, Biological Conservation, 137, 424-436. https://doi.org/10.1016/j.biocon.2007.02.020
Gaoué, O. G., Ticktin T., (2009). Fulani Knowledge of the Ecological Impacts of Khaya Senegalensis (Meliaceae) Foliage Harvest in Benin and its Implications for Sustainable Harvest. Economic Botany, 63, 256-270. https://doi.org/10.1007/s12231-009-9091-6
Gnon, A., (1967). Aménagement de l'espace en pays Bassar : kabou et sa région, FLESH, caen, 160.
Hall, J. B., Aebischer, D. P., Thomlinson, H. F., Osei-Amaning, E., and Hindle, J. R., (1996). Vitellaria Paradoxa a Monograph. Bangor, Royaume-Uni, School of Agriculture and Forest Sciences, University of Wales, 105.
Kelly, B. A., Bouvet, J.-M., and Picard, N. (2004). Size Class Distribution and Spatial Pattern of Vitellaria Paradoxa in Relation to Farmers' Practices in Mali. Agroforestry Systems, 60 (1), 3-11. https://doi.org/10.1023/B:AGFO.0000009400.24606.e3
Koulibaly, A., Goetze, D., Porembski, S., Traoré, D., (2010). Modes d'utilisation des terres et biodiversité : la production agricole et agrobiodiversité. Konaté S and Kampmann D (eds). 2010 : Atlas de la Biodiversité de l'Afrique de l'Ouest, Tome III : Côte d'Ivoire. Abidjan and Frankfurt/Main, 350-361.
Ky-Dembele, C., Tigabu, M., Bayala, J., Ouédraogo, S. J., and Odén, C. P., (2007). The Relative Importance of Different Regeneration Mechanisms in a Selectively Cut Savanna-Woodland in Burkina Faso, West Africa. Forest Ecology and Management, 243(1), 28-38. https://doi.org/10.1016/j.foreco.2007.01.091
Maingi, J. K., Marsh, S. E., (2006). Composition, Structure, And Regeneration Patterns in a Gallery Forest Along the Tana River Near Bura, Kenya, Forest Ecology and Management 236, 211-228. https://doi.org/10.1016/j.foreco.2006.09.006
Mama, A., Bamba, I., Sinsin, B., Bogaert, J., and De Cannière, C., (2014). Déforestation, savanisation et développement agricole des paysages de savanes-forêts dans la zone soudano-guinéenne du Bénin. Bois et Forêts des Tropiques, 324(4),65-75. https://doi.org/10.19182/bft2014.322.a31237
Nacoulma, B. M. I., Traoré, S., Hahn, K., and Thiombiano, A. (2011). Impact of Land Use Types on Population Structure and Extent of Bark and Foliage Harvest of Afzelia Africana and Pterocar- Pus Erinaceus in Eastern Burkina Faso, International Journal Of Biodiversity and Conservation, 3(3), 62-72.
Okullo, J. B. L., Hall, B. J., and Obua, J.(2004). Leafing, Flowering and Fruiting of Vitellaria Paradoxa Subsp. Nilotica in Savanna Parklands in Uganda. Agroferstry Systems, 60, 77-91. https://doi.org/10.1023/B:AGFO.0000009407.63892.99
Ouédraogo, A., Thiombiano, A., Hahn-Hadjali, K., and Guinko, S. (2006). Structure du peuplement juvénile et potentialités de régénération des ligneux dans l'Est du Burkina Faso. Études de la Flore et de la Végétation du Burkina Faso, 10, 17-24.
Ouédraogo, A., Thiombiano, A., Hahn-Hadjali, K., and Guinko, S., (2009). Diversité et dynamique de la végétation ligneuse juvénile du Parc national d'Arly (Burkina Faso). Candollea, 64, 257-278.
Potapou, G. K., (1997). Les facteurs humains et l'évolution de la végétation à Bassar au Nord-Togo. Mémoire de maîtrise, Géographie, Université du Bénin, 96.
RGPH4, (2010). Quatrième Recensement Général de la Population et de l'Habitation du Togo, 29.
Robert, A. (2003). Simulation of the Effect of Topography and Tree Falls on Stand Dynamics and Stand Structure of Tropical Forests. Ecological Modelling, 167, 287-303. https://doi.org/10.1016/S0304-3800(03)00200-X
Ræbild, A., Hansen U. B., and Kambou S., (2012). Regeneration of Vitellaria Paradoxa and Parkia Biglobosa. Agroforest Syst, in a Parkland in Southern Burkina Faso, 85. https://doi.org/10.1007/s10457-011-9397-0
Schumann, K., Wittig, R., Thiombiano, A., Becker, U., and Hahn, K., (2011). Impact of Land-Use Type and Harvesting on Population Structure of a Non-Timber Forest Product-Providing Tree in a Semi-Arid Savanna, West Africa. Biological Conservation, 144, 2369-2376. https://doi.org/10.1016/j.biocon.2011.06.018
Sinsin, B., Eyogmatig, O., Assogbadjo, A. E., Gaoué, O. G., and Sinadouwirou, T. (2004). Dendrométric Characteristics as Indicator of Pressure of Afzelia Africana Sm. Trees Dynamics in Different Climatic Zones of Benin, Biodiversity Conservation, 13, 1555-1570. https://doi.org/10.1023/B:BIOC.0000021328.56517.46
 This work is licensed under a: Creative Commons Attribution 4.0 International License
This work is licensed under a: Creative Commons Attribution 4.0 International License
© Granthaalayah 2014-2022. All Rights Reserved.

